|
| Not quite what I had in mind |
As an avid not-birder I found myself out of my depth and so enlisted the help of an expert; Chris Tessigila-Hymes of the Cornell Lab of Ornithology who graciously agreed to help me out. Chris is an skilled birder and fellow acoustics nut. He is also the listowner of three birding eList list serves, including one dedicated night flight calls: the NFC-L eList. For the record, anything that sounds relatively intelligent and accurate in this post may be attributed to Chris. Any flubs are entirely the fault of the author.
 |
| Chris Tessagila-Hymes (Ornithologist extremus). Identifying features: bird-friendly coffee (for the early bird), birder t-shirt (to avoid confusion with mycologists) and beard (doubles as a bird hide and insulation) |
Night flight calls, Chris tells me, are used to describe the zeeps, chips, soft whistles and other calls that are produced by migrating birds. Like birdsong these calls may be used to identify species or group of species. However, night flight calls differ greatly in their structure, being only milliseconds in duration and orders of magnitude quieter than song. Moreover, where song is produced primarily by males and used as a territorial display, it is thought that both sexes produce night flight calls during the spring and fall migration and the purpose of the call is much more nuanced.
Each year 3-5 billion birds make the round trip migration to southern wintering grounds. Some birds, such as the arctic tern, travel from the arctic to the arctic to antarctic twice a year. These birds have evolved to carefully take advantage of abundant food resources at specific regions throughout their journeys. Therefore it is critical to identify and protect key habitat along the migratory routes in order to ensure the success of these species. Unfortunately, it can be prohibitively expensive to to study migratory routes as scientists have historically been restricted to using expensive technology such as RADAR or geolocators. However, over the last few decades recording technology has become increasingly affordable, allowing both the novice and expert to gain a better understanding of the migrating birds overhead.
Chris has been recording calls for several seasons.
 |
| One reason small birds may migrate at night |
But birds that opt to migrate at night face additional challenges. Principle among these is how to keep the flock together. When vision fails, sound is often a good alternative means of communication. Night flight calls may serve as contact calls, where members call and listen for others in order to keep the flocks together. The calls could also be used to avoid mid-air collisions or allow birds to form night-migrating flocks that say together in the day to forage. Another hypothesis being considered is that some birds may use night flight calls as a rudimentary form of echolocation. During periods of low visibility, e.g. during the pre-dawn decent into forest or scrub (or central park) the echos from the calls may provide some information about their surroundings.
Given all the potential benefits of producing calls, it would make sense that all migrating birds would produce them. Not so! As you will recall, there are plenty of nighttime predators as well. Owls, most notably have amazing hearing worthy of a post of their own. This is one more reason night flight calls are generally shorter and quieter than birdsong. Interestingly, some migrating birds cheat in the game of evolution; they remain silent but form mixed flocks with species that do produce night flight calls. Bastards.
Anwyho, one of the coolest things about night flight calls is the ease at which novices can get involved. Given his extensive experience I asked him what would be the best way to learn to identify different species of birds at night.
If you have good hearing, you don't need a super setup to listen. Pay attention to the weather and known when to step outside at night to listen. After the passage of a cold front in the fall and after the passage of a warm front in the spring (or before a cold front passes). These are the times when the wind is favorable to each migration season. Here's a useful link to a regularly updated Wind Map.
Also, pay attention to nighttime weather by watching the reflectivity at various NEXRAD Radar stations across the United States. If you see lots of active NEXRAD Radar reflection at night near your listening site, this means birds, insects and bats are aloft! Here are a couple of NEXRAD Radar sites that are useful tools to use: NOAA National Mosaic Radar Loop and Paul Hurtado's Radar Archive.
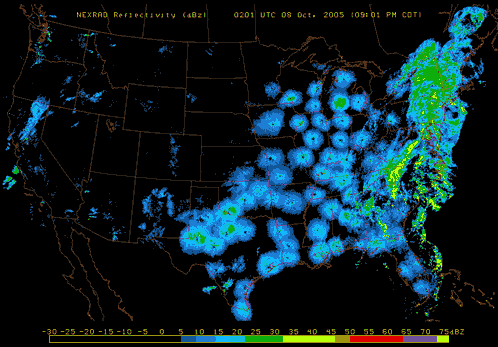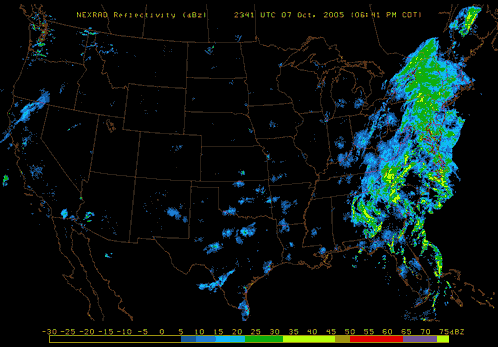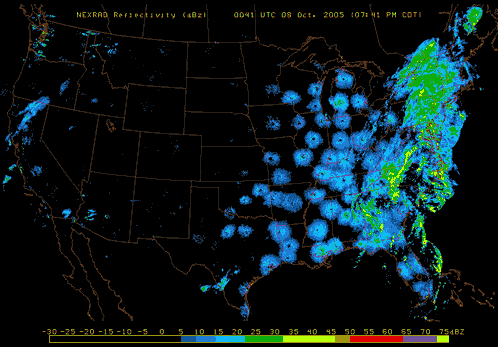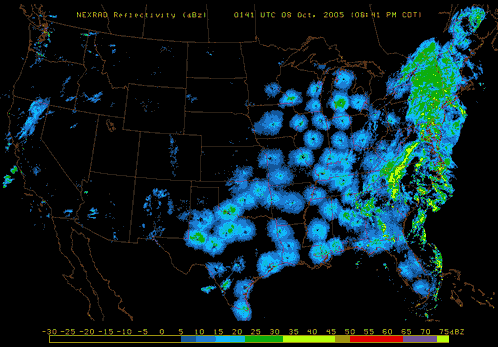 |
| Migrating birds seen taking off at sunset on NASA radar (http://spaceplace.nasa.gov/) |
To build your own recording device Chris suggests checking out Bill Evans' website OldBird.org where instructions are freely available. Several tools area available to analyze collected sound including Raven , Audacity, and Syrinx software. Species identification can be done by comparing recordings to those made by others, e.g. the Evans and O'Brien CD-ROM 'Flight Calls of Migratory Birds' and the night flight library. There are also several detectors on the website but I (the author) have no idea if or how they function so use at your own risk. If you want to discuss your findings with other like minded enthusiasts head over to the to the NFC-L eList. Then, once you are sure of what you've recorded enter your data into the citizen science project eBird following the night flight call reporting protocol.
Okay. If you decide to monitor night migrants with your own rooftop-mounted listening station, I highly recommend you take great care when climbing into and out of a window. During the spring of 2013, I was climbing out my window at night to service the microphone after a rainstorm. As I did this, I somehow awkwardly bent my right knee too much in a lateral direction (the way it's not supposed to bend…). The result of this movement was a perfect "bucket handle" tear of the medial meniscus cartilage in my knee. This required surgical removal of the "handle" part of the tear and effectively rendered me out of commission for the rest of the spring and summer field season that year. Though, happily, I was still quite capable of listening to night migration from my bedroom.
So, kudos to Chris to being more committed to his hobbies than I ever endeavor to be.
Of course a big thanks to you all for reading. Finally, a special, huge, great big thanks to Mr. Chris Tessagila-Hymes for helping out, being a good sport and generally a good person.
Happy Recording!